

I posit that the last universal common ancestor (LUCA) of all life on earth was a lichen, and that this simplifies a lot of the origin of life complexity. Let me explain why.
All life is, of course, descended from one original organism. The last universal common ancestor (aka LUCA) is just that – the last point on a tree that has any surviving branches today. It’s not quite synonymous with “the first living organism”, but it’s probably close, since life differentiates quickly. It’s probably a single-celled microbe, since all of the early cladistic distinctions (bacteria, archaea, protists…) are microbes, so in the hunt for LUCA, we begin in the realm of cellular biology.
“Mainstream” biology will tell you that this LUCA was an extremely minimal critter, probably sort of a cobbled-together organism that was one of the simpler possible collections of biological systems that could be self-sustaining. All complexity of cellular life developed slowly over time from accumulated random mutations.
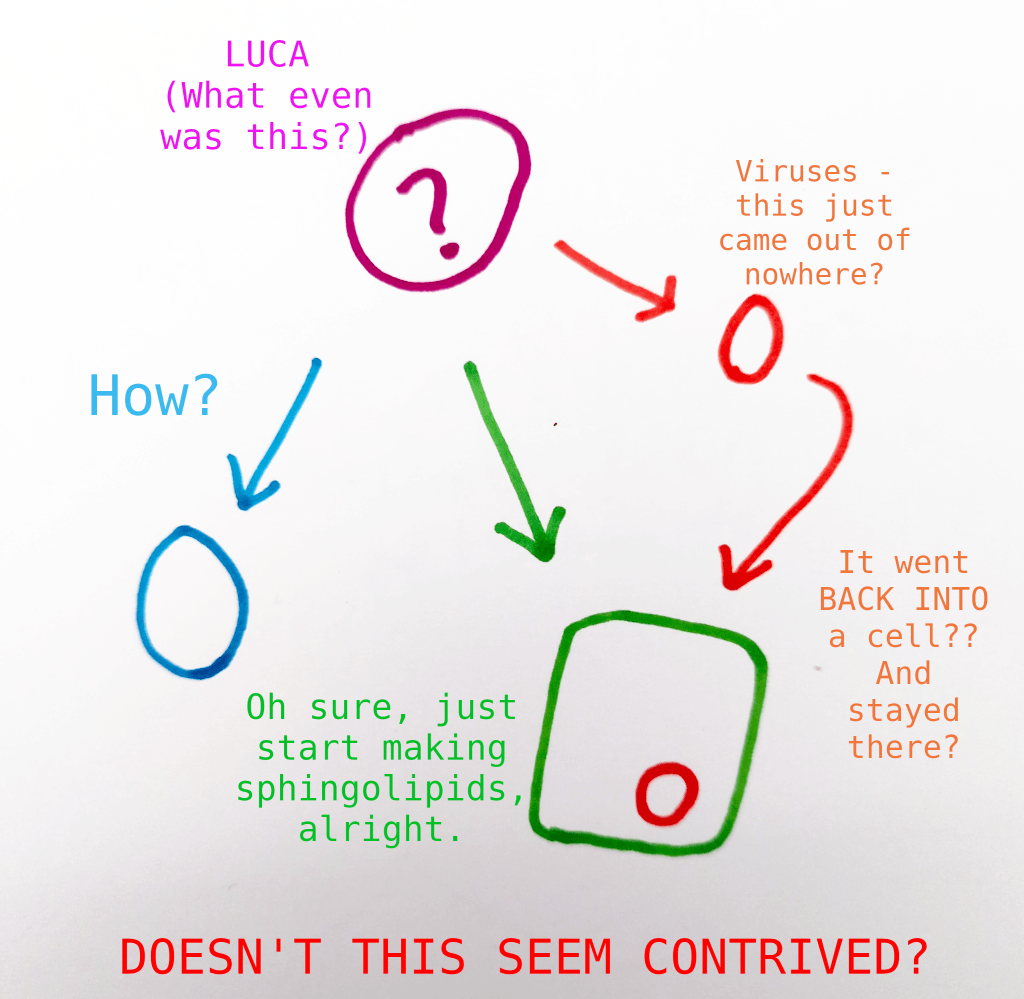
The alternate “Lichen as last universal common ancestor” model, which I will refer to for convenience as “Big Lichen”, does not expect so little of Mother Nature.
Approved scientific dogma describes lichens as a mutual relationship formed between two organisms: a fungi, plus a photosynthetic organism (which is either a eukaryotic algae or a cyanobacteria). The fungi provides structure and substrate with its protective coat, and it feeds off sugars produced by the photosynthetic component. Thus, they team up – or at least this how it is usually seen.
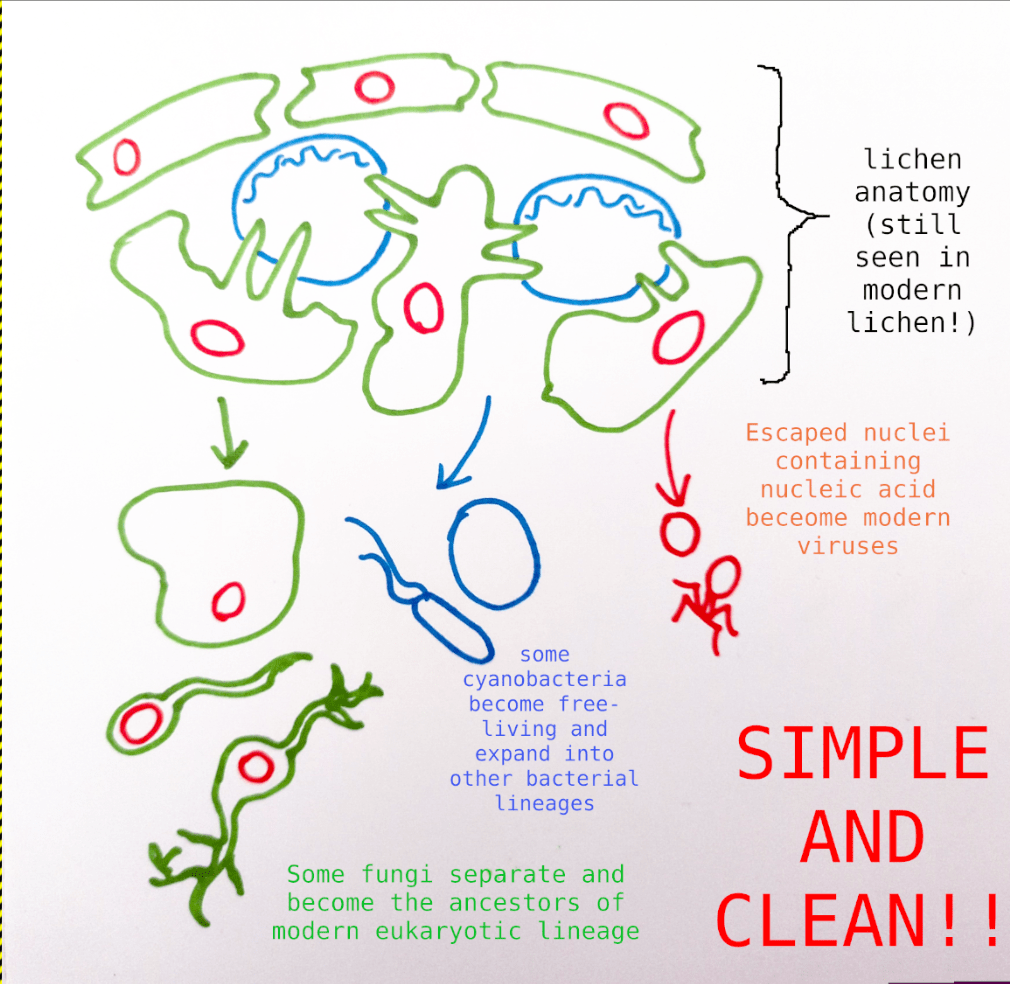
Under this new framework, I believe it makes more sense to view lichens as an entire organism arriving on the evolutionary scene at the right time, and then fungi and cyanobacteria (or algae) on their own as “diminished” or “partial” lichens – and from these, all other lifeforms descended from them, including our own Animalia lineage.
Setting the stage
For right now, let’s be very clear – any abiogenetic explanation for the origin of life requires several deeply unlikely systems to occur at once. The earliest self-sustaining cellular life necessarily requires a replicating genetic material, a metabolism, and some kind of form or structure to sustain the above processes. Without all of these, the others can go no further. This is a problem for any proposal of abiogenesis, not just ours.
Many lichens are self-sustaining systems, however. Once one exists, all it needs is sunlight and a rock to grow on – the one thing we are certain existed in abundance at the dawn of life. Reproduction and dispersal are explained by the lichen’s ability to flake off of rocks and so spread by mechanical action alone, simplifying another step in its early conquest of the earth.
This “lichen earth” would have, over millions of years, contributed biogenic atmospheric gases and rock weathering, slowly building a vast playground of life with ideal conditions for later evolutionary developments and derived forms.
The days between “the origin of life” and “the last universal common ancestor” are, obviously, very fuzzy, because there is no possible genetic evidence form this time period, and early microscopic life left scant fossil evidence. We’ll address in detail the question of how LUCA-as-lichen itself arose later on. For now, how do we get to the modern earth from this lichen-coated paradise?
A more parsimonious start in the garden of life
It’s much, much easier for natural selection to break a system than to build a new one. 99% of random mutations harm the host, rather than help. Many forms which evolution returns to again and again are simplifications of existing structures rather than adding complexity – streamlined wormlike bodyplans have evolved over and over again, for instance, and that’s in a world where worms already exist. Parasites, often the most simple organisms (since they are able to outsource crucial functions to the host), have independently evolved thousands of times.
Even “productive” mutations are often the result of a complex system breaking down. One classic example of natural selection in action is, for instance, the UK’s peppered moths evolving to be darker in color, which allowed them to blend in better with the soot-covered buildings of the industrial revolution. This mutation is a new phenotype that clearly benefits the moth, of course, but it appears to be caused on a genetic level by a transposon inserting multiple copies of a developmental gene, E.G., a broken virus that has fallen into the gene and misfired. Humans who are able to drink milk as adults suffer from a critical childhood enzyme failing to correctly turn off once they’re no longer weaning. The gorgeous antlers of deer, and the magnificent spiral horns of the markhor, are descended from bone cancers.

“Loss of function” mutations are a major source of evolutionary novelty, and they’re immensely more common than “gain of function” mutations. We assume the early development of the tree of life is littered with these “loss of function” mutants.
Whereas current theories of abiogenesis require us to assume numerous “productive” adaptations randomly generated totally de novo in the early days of life, “Big Lichen” theory relaxes this assumption – more events in early life history are more explicable as “destructive” mutations from the original lichen state.
Terminology
Obviously, Big Lichen theory and the dualist nature of the lichen-organism conflicts with our current, simplistic system, which describes an organism as containing strictly one germline. But chimera offspring are very common, even in animals.1 And anyway biologists can handle, like, the entire existence of archaea, we can manage this one.
Let us describe an organism as a reproducing body containing at least one genome subject to natural selection. This more equitable definition encompasses more than you might think – lichens, for instance, both basal and those that have lost and then returned to lichen-nature (more on this later.) Most large plants are, in an expansive sense, lichens: many rely on a symbiotic relationship with a fungal partner to survive at all. Even humans have a practically mandatory symbiosis with numerous partner microbes for digestion, immune health, and more.
We hope the rest of this piece will be clear despite the obvious “so is a lichen a single ‘species’ or what” issue, but it’s clear that further work in this area will be needed to furnish more holistic and appropriate vocabulary and concepts required to truly describe the lichen-centric model of taxonomy.
How did life differentiate, under Big Lichen theory?
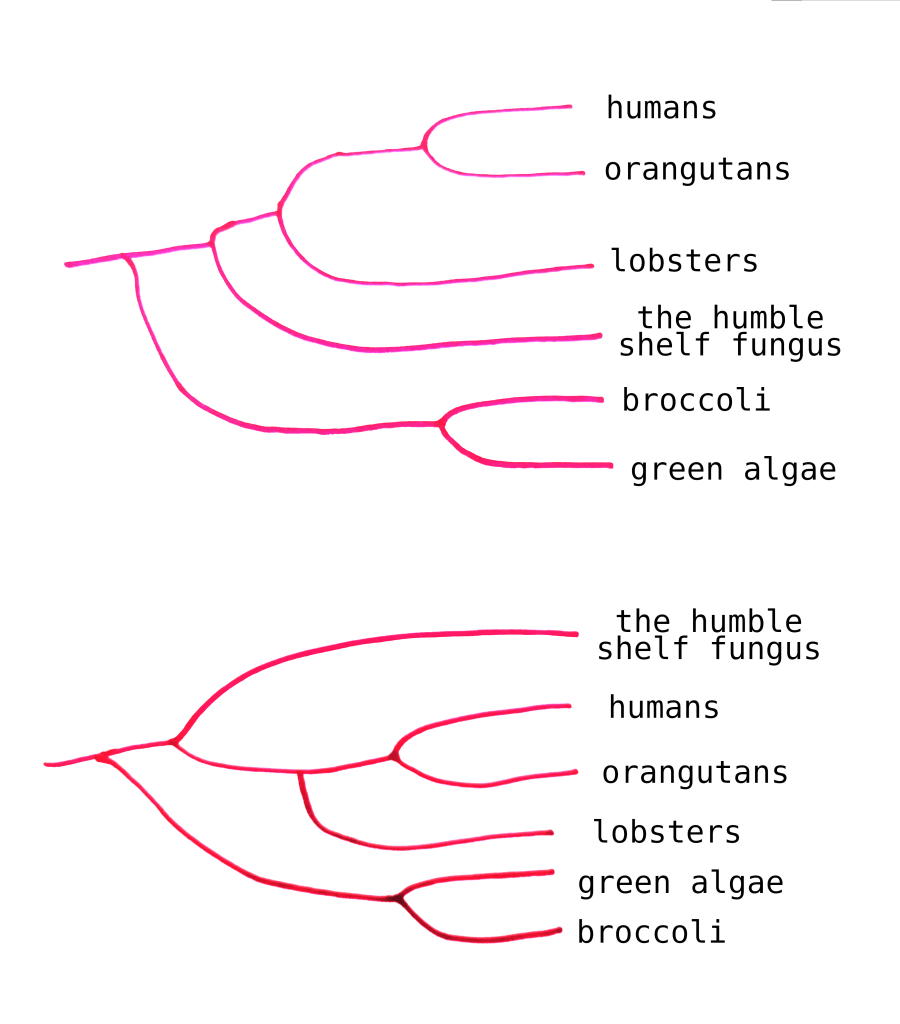
You might know that you can rearrange the specific ordering of a phylogenetic tree. As long as the relationships between the branches stay the same, the order doesn’t matter.
For instance, these two trees convey the same information:
Right? Got that? Okay. We can do a similar procedure to understand the Big Lichen model works. The conventional tree of life looks something like this:
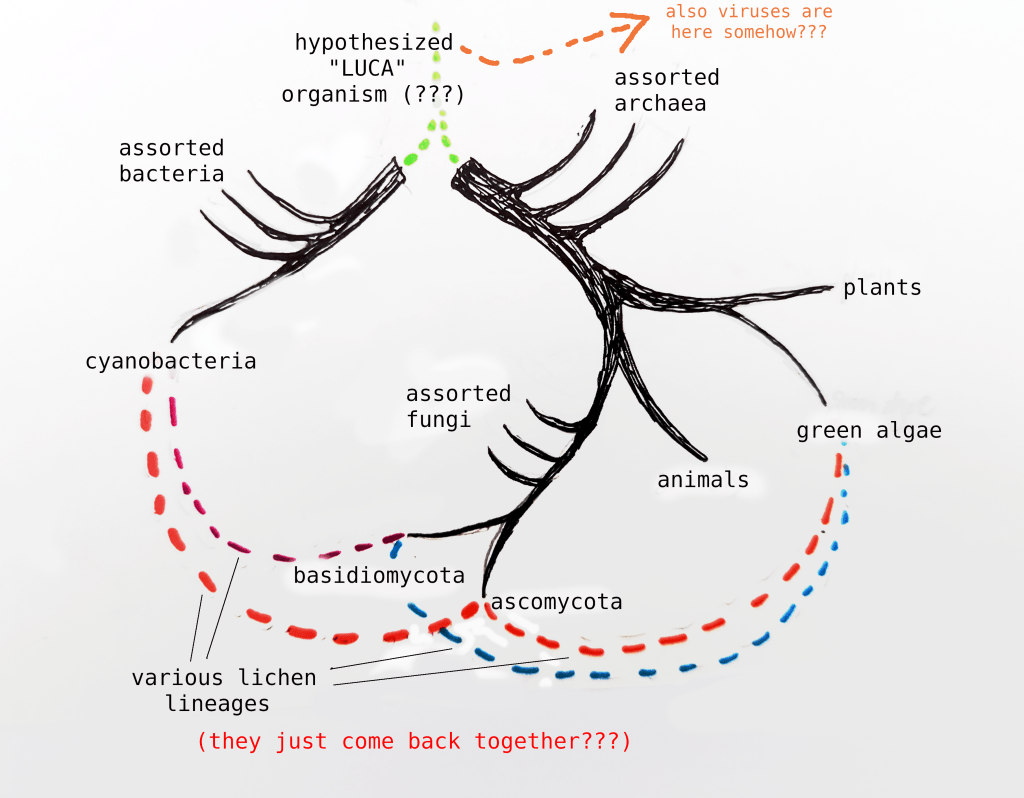
Imagine picking the tree up by one of those lichen lineages. While it’s a little unorthodox, we can actually just slightly rearrange the map to understand the proposed Lichen-As-Common-Ancestor model.
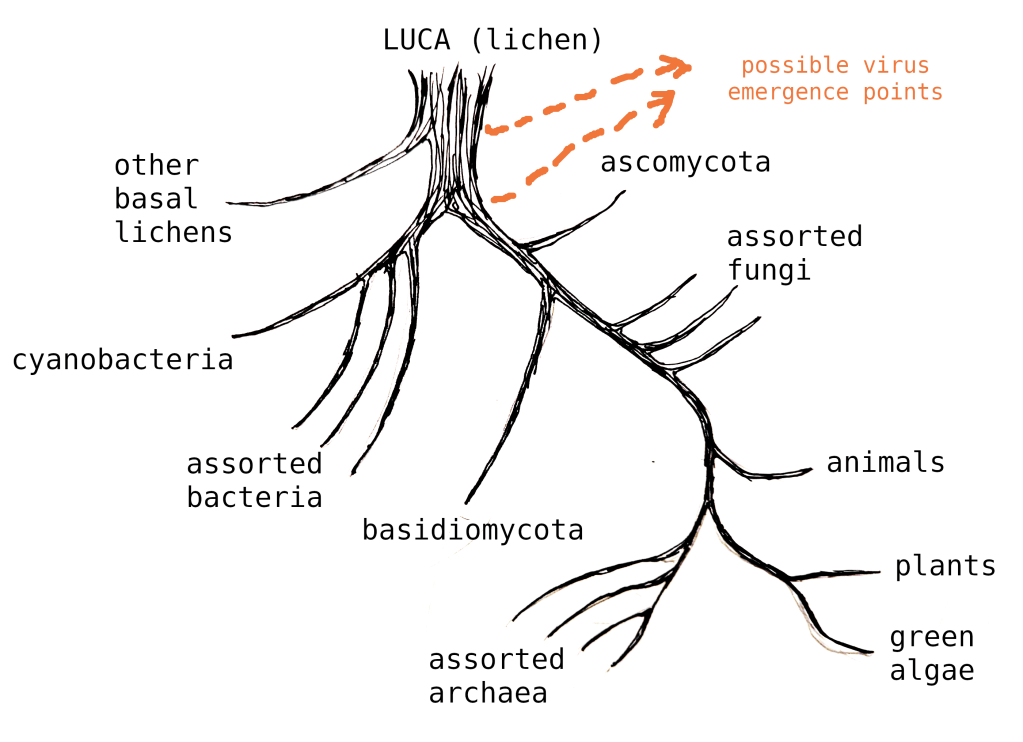
Thus we eliminate the unknown LUCA organism, and cleanly unite the branches.2
What was the ancestral lichen like?
There are multiple kinds of lichen – basidiomycote and ascomycote fungi, with various kinds of algae and cyanobacteria as photosynthetic partners. While it’s not crucial to the theory, it’s worth paying a moment’s thought to what kind of modern lichen the primordial lichen most resembled.
Most modern lichens – 98% – incorporate ascomycota as the fungi partner, and ascomycota are an incredibly diverse clade. They tend to evolve away from being lichens more than they evolve towards being lichens.3 Since all current statistical trends have been true since the beginning of time, we can surely assume that LUCA included an ascomycota.
I will make a more unusual claim regarding the photosynthetic partner. Some 90% of known lichens incorporate a eukaryotic algae, mostly (but not exclusively) green algae. About 10% incorporate a cyanobacteria instead. I believe that cyanobacteria best represent the older and more primordial state of LUCA – for one, bacteria are vastly diverse and ubiquitous, probably the most numerous organisms on earth; suggesting an earlier point of separation from Big Lichen. Also, while bacteria are often thought of as being straightforwardly “simpler” than eukaryotes, they do have several widespread positive characteristics of their own: peptidoglycan cell walls, nitrogen fixation, quorum sensing, etcetra. So I believe we should expect this broad lineage to appear similarly on, with its own traits, alongside its partner eukarytoic fungi.
Under this model, all eukaryotic organisms are descendants of the fungal LUCA lineage. Many developed their own photosynthetic capacity (possibly at some level due to gene exchange with their original partners) and later returned to partnerships with still-present lichenized fungi. This strategy evidently worked and allowed these eukaryote-eukaryote lichen to spread far and wide.
But where did the first lichen come from?
Sandberg et al (2018) claims that the original development of life is so unlikely as to be at the root of the Fermi Paradox – the reason we don’t see signs of alien life, among all the stars of the night sky, is that life is so unlikely to ever spontaneously develop on a random planet that we really shouldn’t expect it to happen more than once per universe (if at all).4
Big Lichen posits an even more complex early organism. Of course, we don’t know where Big Lichen came from – perhaps there was a vast and diverse evolutionary history before it, of which Big Lichen was the only survivor, and thus came forth all extant life. But I want to defend the remote possibility that the first organism ever to exist at all was some form of lichen.
After all, as mentioned repeatedly, the first living organism was already phenomenally complicated. Another relevant concept is the anthropic principle – the fact that sometimes, the fact that you can observe something at all is itself a relevant piece of information. If it takes lichen to eventually form intelligent life, then the only beings that ever perform this analysis and try to find their roots will have come from the tiny set of universes where lichen developed from nothing.
Further evidence comes from the ALNATS statistical method5. Lichen is, of course, very complex, perhaps a million times more complex then the simplest possible first lifeform. For instance, let’s say that the odds of evolutionarily-capable life developing on a suitable planet are 1 in 10,000,000, and the odds of an entire self-sustaining lichen-type organism with two symbiont partners and all developing on a suitable planet are 1 in 10,000,000,000,000. While one looks astonishingly more likely at first, an ALNATS analysis suggest we really shouldn’t worry about this difference.
Is this all a bit abstract? I’ll try to ground it out. There’s a popular creationist argument about the origin of life that goes like this: imagine you’re walking in the desert and you find a pocket watch in the sand. You note its clear numbers, its useful shape and function, its intricate construction. Is it more plausible that the forces of nature have miraculously refined and eroded rocks to result in this intricate shape and situation, or that someone made it?
And ultimately, I do believe that the evidence for abiogenesis is much stronger than the evidence for god, but I see the point. All theories of life require a bunch of bullshit to happen in order to lead to something intricate and without known precedent taking form. And the added layer of complexity does mean Big Lichen is weak evidence for intelligent origins of life.

But we need not abandon abiogenesis. The watch analogy is good. Of course, it’s phenomenally unlikely that lava and metal deposits and desert winds sculpted a working pocket watch, and it’s phenomenally unlikely that the universe created a minimally viable reproducing system of biological molecules, in pools or deep-sea vents of odd organic chemicals. But the universe is big and life is self-reproducing. It only had to happen once.
Now, in your mind’s eye, just replace that pocket watch with a fully functional television with integrated blu-ray player, currently playing a blu-ray copy of Muppet Treasure Island (1996). All around, the lone and level sands stretch far away. Is it more complicated than the pocket watch? Sure. Is it more unlikely than the pocket watch? Sure. But remember: it only has to happen once.
- See, for instance, microchimerism. ↩︎
- Vis-a-vis viruses, Big Lichen theory indicates that viral eukaryogenesis is correct, except that it went in the other direction. ↩︎
- Lutzoni, F., Pagel, M., & Reeb, V. (2001). Major fungal lineages are derived from lichen symbiotic ancestors. Nature, 411(6840), 937-940. ↩︎
- Sandberg, A., Drexler, E., & Ord, T. (2018). Dissolving the Fermi paradox. arXiv preprint arXiv:1806.02404. ↩︎
- “All Long Numbers Are The Same.” ↩︎
